
 Торможение - особый нервный процесс, который обусловливается возбуждением и внешне проявляется угнетением другого возбуждения. Оно способно активно распространяться нервной клеткой и ее отростками. Основал учение о центральноv торможение И. М. Сеченов (1863), который заметил, что изгибающий рефлекс лягушки тормозится при химическом раздражении среднего мозга. Торможение играет важную роль в деятельности ЦНС, а именно: в координации рефлексов; в поведении человека и животных; в регуляции деятельности внутренних органов и систем; в осуществлении защитной функции нервных клеток.
Торможение - особый нервный процесс, который обусловливается возбуждением и внешне проявляется угнетением другого возбуждения. Оно способно активно распространяться нервной клеткой и ее отростками. Основал учение о центральноv торможение И. М. Сеченов (1863), который заметил, что изгибающий рефлекс лягушки тормозится при химическом раздражении среднего мозга. Торможение играет важную роль в деятельности ЦНС, а именно: в координации рефлексов; в поведении человека и животных; в регуляции деятельности внутренних органов и систем; в осуществлении защитной функции нервных клеток.
Типы торможения в ЦНС
Центральное торможение распределяется по локализации на пре-и постсинаптическое;
по характеру поляризации (зарядом мембраны) - на гипер-и деполяризации;
по строению тормозных нейронных цепей - на реципрокное, или соединенное, обратное и латеральное.
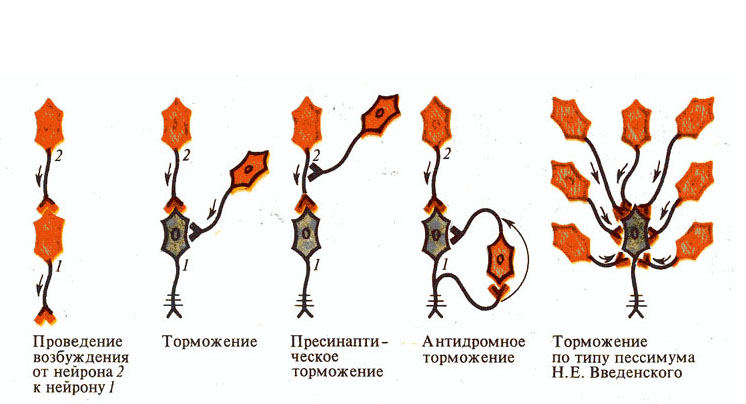
Пресинаптическое торможение, как свидетельствует название, локализуется в пресинаптических элементах и связано с угнетением проведения нервных импульсов в аксональных (пресинаптических) окончаниях. Гистологическим субстратом такого торможения является аксональные синапсы. К возбуждающему аксону подходит вставной тормозной аксон, который выделяет тормозной медиатор ГАМК. Этот медиатор действует на постсинаптическую мембрану, которая является мембраной возбуждающего аксона, и вызывает в ней деполяризацию. Возникшая деполяризация тормозит вход Са2 + из синаптической щели в заключение возбуждающего аксона и таким образом приводит к снижению выброса возбуждающего медиатора в синаптическую щель, торможение реакции. Пресинаптическое торможение достигает максимума через 15-20 мс и длится около 150 мс, то есть гораздо дольше, чем постсинаптическое торможение. Пресинаптическое торможение блокируется судорожными ядами - бикулином и пикротоксин, которые являются конкурентными антагонистами ГАМК.
Постсинаптическое торможение (ГПСП) обусловлено выделением пресинаптическим окончанием аксона тормозного медиатора, который снижает или тормозит возбудимость мембран сомы и дендритов нервной клетки, с которой он контактирует. Оно связано с существованием тормозных нейронов, аксоны которых образуют на соме и дендритах клеток нервных окончаний, выделяя тормозные медиаторы - ГАМК и глицин. Под влиянием этих медиаторов возникает торможение возбуждающих нейронов. Примерами тормозных нейронов являются клетки Реншоу в спинном мозге, нейроны грушевидные (клетки Пуркинье мозжечка), звездчатые клетки коры большого, мозга и др..
Исследованием П. Г. Костюка (1977) доказано, что постсинаптического торможения связано с первичной гиперполяризацией мембраны сомы нейрона, в основе которой лежит повышение проницаемости постсинаптической мембраны для К +. Вследствие гиперполяризации уровень мембранного потенциала удаляется от критического (порогового) уровня. То есть происходит его увеличение - гиперполяризация. Это приводит к торможению нейрона. Такой вид торможения называется гиперполяризационным.
Амплитуда и полярность ГПСП зависят от исходного уровня мембранного потенциала самого нейрона. Механизм этого явления связан с Сl+. С началом развития ТПСП Сl- входит в клетку. Когда в клетке становится его больше, чем снаружи, глицин конформирует мембрану и через открытые ее отверстия Сl+ выходит из клетки. В ней уменьшается количество отрицательных зарядов, развивается деполяризация. Такой вид торможения называется деполяризационным.
Постсинаптическое торможение локальное. Развивается оно градуально, способное к суммации, не оставляет после себя рефрактерности. Является более оперативным, четко адресованным и универсальным тормозным механизмом. По своей сути это «центральное торможение», которое было описано в свое время Ch. S. Sherrington (1906).
В зависимости от структуры тормозного нейронного цепочки, различают следующие формы постсинаптического торможения: реципрокное, обратное и латеральное, которое является собственно разновидностью обратного.
Реципрокное (сочетанное) торможение характеризуется тем, что в том случае, когда при активизации афферентов возбуждаются, например, мотонейроны мышц-сгибателей, то одновременно (на этой стороне) тормозятся мотонейроны мышц-разгибателей, действующие на этот же сустав. Происходит это потому, что афференты от мышечных веретен образуют возбуждающие синапсы на мотонейронах мышц-агонистов, а через посредство вставного тормозного нейрона - тормозные синапсы на мотонейронах мышц-антагонистов. С физиологической точки зрения такое торможение очень выгодно, поскольку облегчает движение сустава «автоматически», без дополнительного произвольного или непроизвольного контроля.
Обратное торможение. В этом случае от аксонов мотонейрона отходит одна или несколько коллатералей, которые направляются в вставных тормозных нейронов, например, клеток Реншоу. В свою очередь, клетки Реншоу образуют тормозные синапсы на мотонейроны. В случае возбуждения мотонейрона активизируются и клетки Реншоу, вследствие чего происходит гиперполяризация мембраны мотонейрона и тормозится его деятельность. Чем больше возбуждается мотонейрон, тем больше ощутимые тормозные влияния через клетки Реншоу. Таким образом, обратное постсинаптическое торможение функционирует по принципу отрицательной обратной связи. Есть предположение, что этот вид торможения требуется для саморегуляции возбуждения нейронов, а также для предотвращения их перевозбуждению и судорожным реакциям.
Латеральное торможение. Тормозная цепь нейронов характеризуется тем, что вставные тормозные нейроны влияют не только на воспаленную клетку, но и на соседние нейроны, в которых возбуждение является слабым или вовсе отсутствует. Такое торможение называется латеральным, поскольку участок торможения, который образуется, содержится сбоку (латерально) от возбужденного нейрона. Оно играет особенно важную роль в сенсорных системах, создавая явление контраста.
Постсинаптическое торможения преимущественно легко снимается при введении стрихнина, который конкурирует с тормозным медиатором (глицином) на постсинаптической мембране. Столбнячный токсин также подавляет постсинаптическое торможение, нарушая высвобождение медиатора из тормозных пресинаптических окончаний. Поэтому введение стрихнина или столбнячного токсина сопровождается судорогами, которые возникают вследствии резкого усиления процесса возбуждения в ЦНС, в частности, мотонейронов.
В связи с раскрытием ионных механизмов постсинаптического торможения появилась возможность и для объяснения механизма действия Вr. Натриq бромид в оптимальных дозах широко применяется в клинической практике как седативное (успокоительное) средство. Доказано, что такой эффект натрия бромида связан с усилением постсинаптического торможения в ЦНС. -
Роль различных видов центрального торможения
Главная роль центрального торможения заключается в том, чтобы во взаимодействии с центральным возбуждением обеспечивать возможность анализа и синтеза в ЦНС нервных сигналов, а следовательно, возможность согласования всех функций организма между собой и с окружающей средой. Эту роль центрального торможения называют координационной. Некоторые виды центрального торможения выполняют не только координационную, а и защитную (охранную) роль. Предполагают, что основная координационная роль пресинаптического торможения заключается в угнетении в ЦНС малосущественными афферентными сигналами. За счет прямого постсинаптического торможения согласуется деятельность антагонистических центров. Обратное торможение, ограничивая максимально возможную частоту разрядов мотонейронов спинного мозга, выполняет и координационную роль (согласовывает максимальную частоту разрядов мотонейронов со скоростью сокращения мышечных волокон, которые они иннервируют) и защитную (предотвращает возбуждению мотонейронов). У млекопитающих этот вид торможения распространен в основном в спинномозговых афферентных системах. В высших отделах мозга, а именно в корковом веществе большого мозга, доминирует постсинаптическое торможение.
Какое функциональное значение пресинаптического торможения? За его счет осуществляется воздействие не только на собственный рефлекторный аппарат спинного мозга, но и на синаптические переключения ряда восходящих по головному мозгу трактов. Известно также о нисходящем пресинаптическом торможении первичных афферентных волокон группы Аа и кожных афферентов. В этом случае пресинаптическое торможение является, очевидно, первым «ярусом» активного ограничения информации, поступающей извне. В ЦНС, особенно в спинном мозге, пресинаптическое торможение часто выступает в роли своеобразной отрицательной обратной связи, которая ограничивает афферентную импульсацию при сильных (например, патологических) раздражениях и таким образом отчасти выполняет защитную функцию по отношению спинномозговых и выше расположенных центров.
Функциональные свойства синапсов не являются постоянными. В некоторых условиях эффективность их деятельности может расти или уменьшаться. Обычно при высоких частотах раздражения (несколько сот за 1 с) в течение нескольких секунд или даже минут облегчается синаптическая передача. Это явление получило название синаптической потенциации. Такая синаптическая потенциация может наблюдаться и по окончании тетанической стимуляции. Тогда она будет называться посттетанической потенциацией (ПТП). В основе ПТП (долговременного увеличения эффективности связи между нейронами), вполне вероятно, лежат изменения функциональных возможностей пресинаптического волокна, а именно его гиперполяризация. В свою очередь, это сопровождается повышением выхода медиатора в синаптическую щель и появлением увеличенного ВПСП в постсинаптической структуре. Есть данные и о структурных изменениях при ПТП (набухание и рост пресинаптических окончаний, сужение синаптической щели т.д.).
ПТП гораздо лучше выражена в высших отделах ЦНС (например, в гиппокампе, пирамидных нейронах коры большого мозга) по сравнению с спинномозговыми нейронами. Наряду с ПТП в синаптическом аппарате может возникать постактивационная депрессия, выражающаяся уменьшением амплитуды ВПСП. Эту депрессию многие исследователи связывают с ослаблением чувствительности к действию медиатора (десенсибилизации) постсинаптической мембраны или различным соотношением затрат и мобилизации медиатора.
С пластичностью синаптических процессов, в частности с ПТП, возможно, связаны формирования новых межнейронных связей в ЦНС и их закрепление, т.е. механизмы обучения и памяти. Вместе с тем следует признать, что пластические свойства центральных синапсов изучены пока недостаточно.